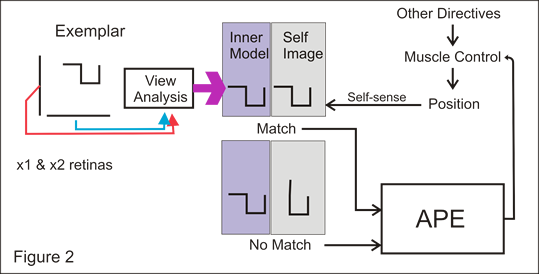
Logic was thought to be the key paradigm for human intelligence from the Aristotle to Wittgenstein, and then in the early days of Artificial Intelligence as implemented on computers.
Careful consideration shows that aping, the ability to copy an action of another individual, is more central to intelligence. It is as much a mammal trait as a human one. Formal exercises in logic are not needed for one animal to learn behavior from another. The complex neural circuitry created by evolutionary pressures to enhance aping potential eventually became the basis of human language and its formalized reasoning skills known as logic.
Aping consists of observing (usually visually) what some other individual is doing, and then copying that behavior. Humans do this so naturally that we seldom think about it consciously. Unless trained to do so (by aping others), practicing logic is a much harder skill. Yet logic circuits are easy to implement with electronic parts. Aping another individual requires very complex systems within the brain.
This essay, and the accompanying computer programs and physical systems, is an attempt to create machines capable of aping, which then should be able to handle other complex behaviors usually associated with human intelligence, including language, mechanical skills, reasoning, and possibly consciousness.
The advantage of aping skills goes beyond helping to ensure the survival of an individual animal. With aping a behavior with survival benefits can become cultural, spanning an unlimited number of generations of animals. This without any genetic change specific to the behavior itself. A generalized pool of neurons can become capable of generating a wide and flexible variety of behaviors, giving a species a heavy advantage over animals that can only engage in behavior based on pre-programmed neural systems.
Pre-language example. Consider a simple aping process and how it differs from paradigms like ordinary remembered (learned) behavior. An animal, a young wolf, has always simply crossed a cold stream by simply plunging through it. One day it sees an older wolf stop at the stream and proceed along the bank to a point where there is a rock in the center of the stream. The older wolf jumps to the rock, then to the far bank, thereby avoiding getting wet while crossing. The young wolf then apes that behavior. Afterwards, when crossing the stream in that area (when not in hot pursuit of prey) the young wolf diverts to the special crossing point.
Note how the aping behavior differs from what might be other reasons for using the rock to cross. The young wolf could learn of the rock crossing by exploration, then would remember the location of the crossing when appropriate. You could argue that following an older animal might be an instinct, so this is not a real example of aping, just a side effect of following.
Aping behavior might have originated in following behavior, and the basic ability to learn from experience, but as more complex situations are involved, it becomes clear that aping is a special case requiring special capabilities.
Supposing this simple example constitutes a form of aping, what can we say about how it is accomplished by the wolf brain? The wolf is capable of recognizing an externalized self. The example wolf, to the aping wolf, represents a possibility for itself. Metaphorically, the aping wolf can see itself crossing the stream using the rock when it sees the example wolf. So the wolf-brain is capable of contemplating (in a very simple, non-philosophic way) the future. It can choose to ape the example, or it can refuse to cross the stream, or it can ignore the example and get wet and cold crossing the stream.
Aping, in the infants of actual apes including humans, is probably automatic. Only at a later time will the young apes start choosing whether or not to ape particular behaviors. There is typically outside support for aping in human children. We reward them when we like their aping efforts, but may discourage them when we don't like their efforts (as when they engage in behaviors culturally reserved for adults).
Consider the "simple" modern act of making a piece of toast with a bread toasting appliance. A child is not likely to learn to do this by accidentally taking a slice of bread, sticking it into the appliance, and pushing down the lever. A child learns this by watching a example or exemplar. The child understands that doing what an exemplar does leads to the same result. The child sees that taking a piece of bread, placing it in a certain way in the appliance, and pressing the lever results in the making of toast. Usually, if sufficiently coordinated, a child can do this on the first try. Aping often results in some level of success in achieving a goal on the first try.
To ape an exemplar the aping child must already have similar levels of muscle control and sense of where its body is in relation to its surroundings. It must understand that aping the exemplar's movements (or other behaviors like vocalizing) achieves a desired result. If an aping attempt is made that does not achieve the desired results, a child may conclude that something is missing from the aping process, a subtlety or trick.
To ape, the brain must have a number of capabilities, which might be summed up as the ability to coordinate the body of the subject with the body of the exemplar.
In humans, when children are quick to ape adults, we call them intelligent. If they are slow to imitate adult behaviors, we call the slow or less intelligent.
Language learning example. Human babies start learning to imitate vocalizations soon after birth. Vocal fussing often results in rewards, like feeding, encouraging further vocalizations. The process may seem like a long one when mechanical devices have long existed that can record and play back sounds and language. There is a fundamental difference between the mechanical play back model and the aping vocalization model that is critical to the reconstruction of how humans are capable of intelligent behavior, including understanding how the world works. [I prefer Machine Understanding as a term for what we are working towards with computerized robots capable of aping, because Artificial Intelligence has largely ignored a set of crucial problems that logic circuits have difficulty handling].
The human brain does not appear to be a recording medium in the sense that a phonograph, magnetic tape machine, or silicon-memory based sound recording and playing device is. The human ability to immediately repeat a phrase of language or hum a melody does not arise from a like mechanism. Humans have fairly good memories, but a weak ability to memorize.
Consider the problem, for the human brain, of learning to repeat its first few spoken words, say "mama, papa, no, puppy, bye, milk." In the ear there are a number of sensory cells that respond to different frequencies of sound. They send nerve impulses towards the brain. At the other end we have the sound making machinery: lungs, vocal cords, and mouth. An infant screaming soon after birth indicates that machinery works, but is nothing like language. Months of hearing and experimental vocalizations follow. There may be feedback, positive or negative, by parents and other exemplars, as well as careful repetition of simple words in the hope this will induce aping by the child.
In the end, however, success is the ability of the child to say a word that sounds, to its own ear, like the kind of words spoken by the exemplars. It will not be identical, nor will it ever be identical, in its fine structure, in fact it will be identifiably different, an individual voice. Six words mastered, many more will follow, all aped after exemplars. In addition, the words will have meaning: they will coordinate with objects or actions in the external world (or internal world, as with "more" or "hurts").
The neural machine that coordinates heard words and the speaking of words with an "ego" must be very complex and very capable. Recreate that machine and we would have a model that, perhaps with some specialized variations, would be able to ape visual input to body positions, or anything else required of it.